

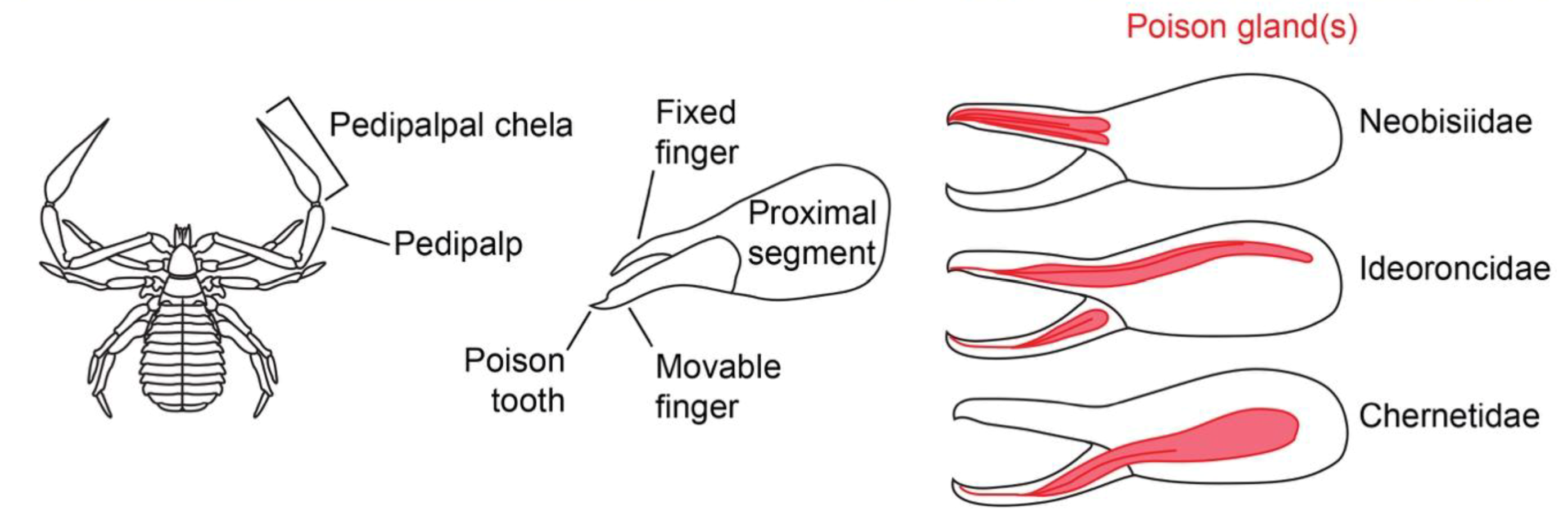
A pseudoscorpion (family Chernetidae) scurries away from daylight, eventually tucking its left pincer in and disappearing between the layers of bark. Pseudoscorpions are a fascinating order of arachnids, comprised of over 3,000 species averaging just 2–4 millimeters in length as adults. Despite their resemblance to scorpions, recent phylogenetic analyses place pseudoscorpions as the outgroup to scorpions, uropygi, amblypygi, and spiders. In addition, pseudoscorpions have a number of striking anatomical distinctions: (1) the absence of book lungs and respiration through tracheae and abdominal spiracles, (2) most notably, the lack of modified metasomal segments into a caudal stinger, (3) the presence of spinnerets at the tips of the chelicerae, and (4) a venom apparatus at the tip of the chelate pedipalps. Not much work has been done in the sensory ecology of pseudoscorpions, though many accounts attribute a chemosensory and mechanosensory function to the fine hairs of the pincer-modified pedipalps. This could explain their locomotor patterns of moving around with the claws fully extended, positioned just above the ground like a metal detector.

Pseudoscorpions are most well known for their phoretic behavior; a major component of their life histories involves hitchhiking on larger arthropods. As opposed to mites which simply latch on with fine tarsal claws, pseudoscorpions use one of their pincer-modified pedipalps to tightly grasp the exoskeleton of their host, most often being the legs. Phoresy in pseudoscorpions has been documented in numerous arthropod orders, including beetles, flies, wasps, butterflies, grasshoppers, caddisflies, hemipterans, dragonflies, neuropterans, spiders, and opiliones (just to name a handful I came across in the literature). There is even evidence of hitchhiking on bats and foraging in the nests of packrats! Pseudoscorpions have one of two types of symbiotic relationships with their hosts: (1) commensalism; only the pseudoscorpion benefits from enhanced dispersal ability, and (2) mutualism, in which pseudoscorpion consumption of parasitic larvae and mites benefits its phoretic host.

Of the more famous pseudoscorpion species, there is Cordylochernes scorpiodes (Chernetidae), a phoretic specialist of the neotropical harlequin beetle (Acrocinus longimanus). Female Cordylochernes incubate eggs and deposit nymphs on the same decaying trees (e.g. Ficus) which offer food resources and laying sites for harlequin beetles. Decaying trees are an ephemeral resource in tropical forests, creating a strong selective pressure for the pseudoscorpions to locate newly-fallen trees. As a solution to this dispersal conundrum, there is evidence that Cordylochernes can assess the quality of trees, boarding harlequin beetles when residing on older, resource-depleted trees, and disembarking on newly fallen trees.

By using harlequin beetles for transport, Cordylochernes not only increases the the probability of encountering more suitable habitat, but also the probability of running into potential mates. Just as in territory defense, male pseudoscorpions actively compete to defend their phoretic beetle hosts (particularly the abdomen and under the elytra) as strategic locations to encounter and mate with dispersing females. While large male size confers an advantage on the beetle battleground, under the low population density conditions of decaying trees, sexual selection favors smaller males that mature more rapidly. Studies suggest that under this selective regime, dispersal has played a role in the evolution of sexual size dimorphism and that fluctuating sexual selection may explain the high phenotypic variation in this species.

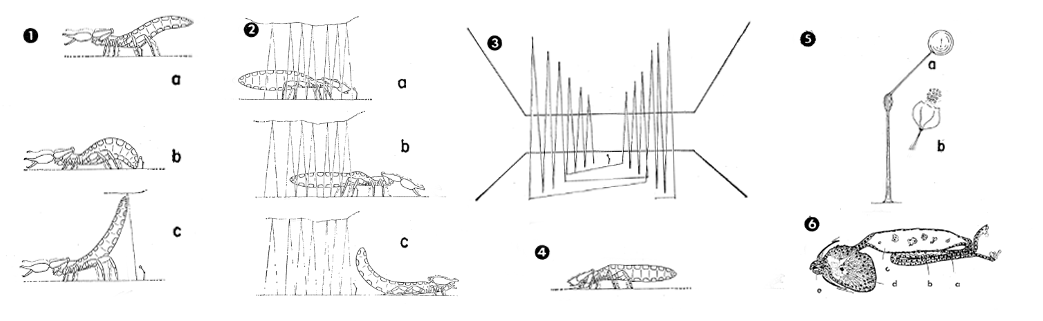
Pseudoscorpions have a variety of courtship behaviors, but they have only been characterized in a few species. In some families (Chernetidae & Cheliferidae), males simply deposit the spermatophore in the absence of the females, relying entirely on the female’s chemotactic ability for localization. In other groups, more complex behaviors have evolved, such as in the olpiid pseudoscorpion, Serianus. First, in the presence of a female, a male shakes his pedipalps in an advance, quickly moving the body back and forth when approaching too closely. The behavior continues until either the female rejects the male, or the male is able to grasp the female’s two pedipalps by his own. This scenario is reminescent of the courtship ritual of true scorpions, though it does not constitute a life-or-death scenario due to the lack of a stinger.
After a short time, the male will turn his attention away from the female and search erratically for a suitable deposition site, between two layers of wood of a particular distance apart. He will then deposit the spermatophore, connected to the substrate by a thin silken stalk just a fifth of a centimeter high, and quickly mark a maze-like array of threads that tapers into the spermatophore location. In contrast to other pseudscorpions, Serianus possesses an additional opisthosomal silk gland which is exclusively used in path formation to the spermatophore. This is distinct from the spinnerets of the chelicerae that is well documented in producing silken nests for overwintering. When a receptive female locates the threaded path (sometimes guided by the male), she will search for the spermatophore with the pedipalps and uptake it into the abdomen. The outer surface of the spermatophore breaks due to hydrostatic pressure, completing the mating ritual.
It is incredible that such complex courtship behaviors reside in such a tiny arthropod. I hope you’ve found this description as fascinating as I do!
This is fascinating. I’ve never seen a pseudoscorpion. I hope to find one eventually. Great photos, as always.
Thank you! Glad you enjoyed the post. If you dig around under logs and debris and look very closely, you may have luck finding one!